
Page 128 - 《广西植物》2026年第3期
P. 128
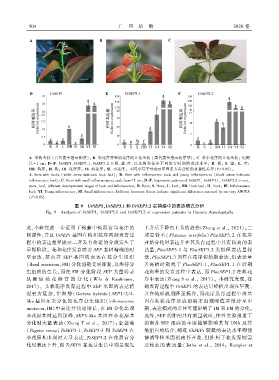
5 0 4 广 西 植 物 2026 卷
A. 带钩茎枝 (白色箭头指示钩芽)ꎻ B. 带花序芽和幼花序的开花茎枝 (黑色箭头指示花序芽)ꎻ C. 带小花序的开花茎枝ꎻ 比例
尺 = 1 cmꎻ D-F. UrSEP1、UrSEP3.1、UrSEP3.2 在根、茎、叶ꎬ以及钩和花序不同发育时期的表达水平ꎻ R. 根ꎻ S. 茎ꎻ L. 叶ꎻ
HB. 钩芽ꎻ H. 钩ꎻ IB. 花序芽ꎻ YI. 幼花序ꎻ SI. 小花序ꎮ 不同小写字母表示单因素方差分析的显著性差异(P<0.05)ꎮ
A. Stem with hooks (white arrow indicates hook bud)ꎻ B. Stem with inflorescence buds and young inflorescences (black arrow indicates
inflorescence bud)ꎻ C. Stem with small inflorescencesꎻ scale bars= 1 cmꎻ D-F. Expression patterns of UrSEP1ꎬ UrSEP3.1ꎬ UrSEP3.2 in rootꎬ
stemꎬ leafꎬ different developmental stages of hook and inflorescenceꎻ R. Rootꎻ S. Stemꎻ L. Leafꎻ HB. Hook budꎻ H. Hookꎻ IB. Inflorescence
budꎻ YI. Young inflorescenceꎻ SI. Small inflorescence. Different lowercase letters indicate significant differences assessed by one ̄way ANOVA
(P<0.05).
图 9 UrSEP1、UrSEP3.1 和 UrSEP3.2 在钩藤中的表达模式分析
Fig. 9 Analysis of UrSEP1ꎬ UrSEP3.1 and UrSEP3.2 of expression patterns in Uncaria rhynchophylla
此ꎬ本研究进一步证明了钩藤中钩器官与花序的 上升后下降再上升的趋势(Zhang et al.ꎬ 2021)ꎻ二
同源性ꎬ并且 UrSEPs 基因在钩和花序两种发育过 球悬铃木( Platanus acerifolia) PlacSEP3.2 在花序
程中的表达差异提示二者发育命运的分歧发生于 开始分化时表达并在其发育过程中具有较高的表
早期阶段ꎮ 花和花序发育需要 SEP 基因精确的时 达量ꎬPlacSEP3.1 与 PlacSEP3.2 类似但表达量较
空表达ꎬ 拟 南 芥 SEP 基 因 的 表 达 在 花 分 生 组 织 低ꎬPlacSEP1.3 同样在花序初始期表达ꎬ但表达量
(floral meristemꎬFM)分化前期受到抑制ꎬ以维持分 升高的时期晚于 PlacSEP3.1ꎬPlacSEP1.1 在后期
生组织的生长ꎬ而在 FM 分化阶段 SEP 大量转录 花和果的发育过程中表达ꎬ而 PlacSEP1.2 在雌花
从 而 启 动 花 器 官 的 分 化 ( Wils & Kaufmannꎬ 序中表达(Zhang S et al.ꎬ 2017)ꎮ 本研究发现ꎬ在
2017)ꎮ 头状花序发育过程中 SEP 基因的表达情 钩发育过程中 UrSEP1 的表达量峰值出现在早期ꎬ
况更为复 杂ꎬ非 洲 菊 ( Gerbera hybrida) SEP1 / 2 / 4 ̄ 并在钩形成期降至低谷ꎬ而花序发育过程中该基
like 基因在未分化的花序分生组织( inflorescence 因在头状花序形成初期才出现峰值并维持至中
meristemꎬIM)中表达并快速增长ꎬ在 IM 分化出现 期ꎬ表达模式的差异可能影响了 IM 和 FM 的分化ꎮ
小花原基时达到顶峰ꎬSEP3 ̄like 基因在小花原基 此外ꎬSEP 的作用具有剂量效应ꎬ体外实验报道了
分化时大量表达( Zhang T et al.ꎬ 2017)ꎻ金盏菊 拟南芥 SEP 蛋白的丰度能够影响其与 DNA 及其
(Tagetes erecta) TeSEP3 ̄1、TeSEP3 ̄3 和 TeSEP4 在 他蛋白的结合ꎬ棉花 GhSEPs 较低的表达水平即能
小花原基出现时大量表达ꎬTeSEP3 ̄2 在花器官分 够诱导转基因拟南芥开花ꎬ但作用于花发育则需
化时表达上升ꎬ而 TeSEP1 在花芽生长中则呈现先 要较高 的 表 达 量 ( Jetha et al.ꎬ 2014ꎻ Rumpler et