
Page 37 - 《广西植物》2020年第7期
P. 37
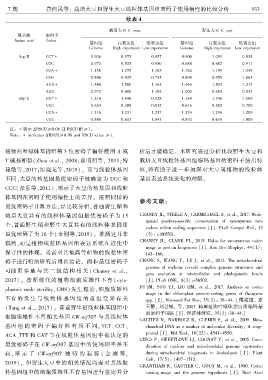
7 期 唐向民等: 栽培大豆和野生大豆线粒体基因组密码子使用偏性的比较分析 9 3 3
续表 4
栽培大豆 G. max 野生大豆 G. soja
氨基酸 密码子
Amino acid Codon
基因组 高表达组 低表达组 基因组 高表达组 低表达组
Genome High experssion Low experssion Genome High experssion Low experssion
Arg / R CGT∗ 0.906 0.975 0.857 0.900 1.091 0.835
CGC 0.672 0.525 0.980 0.684 0.682 0.911
CGA∗ 1.158 1.275 1.163 1.164 1.159 1.139
CGG 0.846 0.825 0.735 0.804 0.955 1.063
AGA∗ 1.446 1.800 1.163 1.446 1.432 1.215
AGG 0.972 0.600 1.102 1.002 0.682 0.835
Gly / G GGT∗ 1.216 1.846 0.928 1.168 1.746 1.100
GGC 0.624 0.308 0.812 0.616 0.282 0.700
GGA∗ 1.316 1.231 1.217 1.324 1.296 1.200
GGG 0.848 0.615 1.043 0.892 0.676 1.000
注: ∗表示 ΔRSCU≥0.08 且 RSCU 值≥1ꎮ
Note: ∗ indicates ΔRSCU≥0.08 and RSCU value ≥1.
植物的叶绿体基因组第 3 位密码子偏好使用 A 或 析后才能确定ꎮ 本研究通过分析比较野生大豆和
T 碱基相似(Zhou et al.ꎬ 2008ꎻ尚明照等ꎬ 2011ꎻ傅 栽培大豆线粒体基因组编码基因的密码子使用特
建敏等ꎬ2017ꎻ原晓龙等ꎬ2019)ꎮ 而与线粒体基因 征ꎬ将有助于进一步加深对大豆属植物的线粒体
不同ꎬ大豆的核基因最优密码子被确定为 UCC 和 基因表达系统进化的理解ꎮ
GCC(张乐等ꎬ2011)ꎬ暗示了大豆的核基因和线粒
体基因在密码子使用偏性上的差异ꎮ 按照同样的
参考文献:
最优密码子计算方法ꎬ经比较分析ꎬ普通野生稻和
栽培大豆共有的线粒体基因组最优密码子为 15 CHANEY JLꎬ STEELE Aꎬ CARMICHAEL Rꎬ et al.ꎬ 2017. Wide ̄
spread position ̄specific conservation of synonymous rare
个ꎬ普通野生稻和野生大豆共有的线粒体基因组
codons within coding sequences [J]. PLoS Comput Biolꎬ 13
最优密码子为 16 个(金刚等ꎬ2019)ꎮ 推测这并非 (5): e1005531.
偶然ꎬ而是植物线粒体基因组表达系统在进化中 CHANEY JLꎬ CLARK PLꎬ 2015. Roles for synonymous codon
usage in protein biogenesis [J]. Ann Rev Biophysꎬ 44(1):
保守性的体现ꎮ 尚需对其他高等植物的线粒体密 143-166.
码子进行归纳研究后得出结论ꎮ 而非最佳密码子 CHANG Sꎬ WANG Yꎬ LU Jꎬ et al.ꎬ 2013. The mitochondrial
genome of soybean reveals complex genome structures and
可能更 多 地 与 其 二 级 结 构 相 关 ( Chaney et al.ꎬ
gene evolution at intercellular and phylogenetic levels
2017)ꎮ 按照现代的植物细胞质雄性不育( cyto ̄ [J]. PLoS ONEꎬ 8(2):e56502.
plasmic male sterilityꎬCMS) 发生理论ꎬ细胞质雄性 FU JMꎬ SUO YJꎬ LIU HMꎬ et al.ꎬ 2017. Analysis on codon
usage in the chloroplast protein ̄coding genes of Diospyros
不育 的 发 生 与 线 粒 体 基 因 组 的 重 组 变 异 有 关 spp. [J]. Nonwood For Resꎬ 35(2): 38-44. [傅建敏ꎬ 索
(Tang et al.ꎬ 2017)ꎮ 普通野生稻线粒体基因组中 玉静ꎬ 刘慧敏ꎬ 等ꎬ 2017. 柿属植物叶绿体蛋白质编码基
因密码子用法 [J]. 经济林研究ꎬ 35(2):38-44.]
细胞质雄性不育相关基因 CW ̄orf307 与其线粒体
GALTIER Nꎬ NABHOLZ Bꎬ GLEMIN Sꎬ et al.ꎬ 2009. Mito ̄
基因 组 的 密 码 子 偏 好 性 有 所 不 同ꎬ TCT、 CCT、 chondrial DNA as a marker of molecular diversity: A reap ̄
ACA、TTT 和 CAT 等在线粒体基因组中被认定的 praisal [J]. Mol Ecolꎬ 18(22): 4541-4550.
GIEGé Pꎬ SWEETLOVE LJꎬ COGNAT Vꎬ et al.ꎬ 2005. Coor ̄
最优密码子在 CW ̄orf307 基因中的使用频率并不
dination of nuclear and mitochondrial genome expression
高ꎬ 暗 示 了 CW ̄orf307 独 特 的 起 源 ( 金 刚 等ꎬ during mitochondrial biogenesis in Arabidopsis [ J]. Plant
Cellꎬ 17(5): 1497-1512.
2019)ꎮ 但野生大豆中的相关情况尚需对其线粒
GRANTHAM Rꎬ GAUTIER Cꎬ GOUY Mꎬ et al.ꎬ 1980. Codon
体基因组中的细胞质雄性不育基因进行鉴定并分 catalog usage and the genome hypothesis [ J]. Nucl Acid