
Page 64 - 《广西植物》2020年第9期
P. 64
9 期 王芬等: 基于高通量测序的都匀地区福鼎大白种茶树根茎叶分析 1 2 7 9
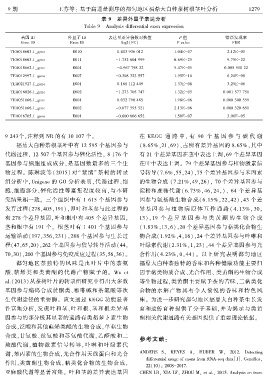
表 9 差异外显子表达分析
Table 9 Analysis differential exon expression
基因 ID 外显子 ID 表达量差异倍数对数值 P 值 错误发现率
Gene ID Exon ID log2(FC) P value FDR
TEA018663.1_gene E010 0.403 936 012 1.04E-07 2.12E-05
TEA018663.1_gene E011 -1.732 604 999 6.69E-25 9.75E-22
TEA018662.1_gene E001 -0.967 798 22 5.47E-05 0.005 510 22
TEA012997.1_gene E007 -0.308 323 557 1.95E-10 6.24E-08
TEA021927.1_gene E001 0.168 112 449 1.33E-08 3.25E-06
TEA016026.1_gene E002 -1.273 705 747 1.32E-05 0.001 577 751
TEA031666.1_gene E001 0.032 790 485 1.98E-06 0.000 300 559
TEA031666.1_gene E002 -0.077 595 321 2.13E-06 0.000 320 651
TEA016705.1_gene E001 -0.600 866 652 1.58E-07 3.06E-05
9 243个ꎬ注释到 NR 的有 10 107 个ꎮ 在 KEGG 通 路 中ꎬ 有 90 个 基 因 参 与 碳 代 谢
福鼎大白种茶根茎叶中有 12 595 个基因参与 (8.65%ꎬ21ꎬ69)ꎬ占所有差异基因的 8.65%ꎬ其中
代谢过程ꎬ 12 507 个基因参与催化活性ꎬ 8 176 个 有 21 个差异基因在茎中表达上调ꎬ69 个差异基因
基因参与细胞组成成分ꎬ是基因数最多的三个生 在叶中表达上调ꎮ 79 个差异基因参与植物激素信
物过程ꎮ 陈琳波等( 2015) 对“ 紫鹃” 茶树的转录 号转导(7.6%ꎬ55ꎬ24)ꎬ75 个差异基因参与苯丙素
组分析中ꎬUnigene 的 GO 分析表明ꎬ代谢过程、细 的生物合成 (7.21%ꎬ49ꎬ26)ꎬ 70 个差异基因参与
胞、细胞部分、催化活性等富集程度较高ꎬ与本研 淀粉和蔗糖代谢(6.73%ꎬ46ꎬ24ꎬ)ꎬ 64 个差异基
究结果相一致ꎮ 三个组织中有 1 615 个基因参与 因参与氨基酸生物合成(6.15%ꎬ22ꎬ42)ꎬ43 个差
发育过程(278ꎬ405ꎬ191)ꎬ即叶和茎参与此过程的 异基 因 参 与 植 物 病 原 体 互 作 通 路 ( 4. 13%ꎬ 30ꎬ
有 278 个差异基因ꎬ叶和根中有 405 个差异基因ꎬ 13)ꎬ19 个 差 异 基 因 参 与 类 黄 酮 的 生 物 合 成
茎和根中有 191 个ꎮ 根茎叶有 1 401 个基因参与 (1.83%ꎬ13ꎬ6)ꎬ20 个差异基因参与萜类化合物生
运输活动(197ꎬ356ꎬ231)ꎬ288 个基因参与生长过 物合成(1.92%ꎬ4ꎬ16)ꎬ24 个差异基因参与卟啉和
程(47ꎬ65ꎬ20)ꎬ262 个基因参与信号转导活动(44ꎬ 叶绿素代谢(2.31%ꎬ1ꎬ23)ꎬ44 个差异基因参与光
70ꎬ30)ꎬ200 个基因参与免疫反应过程(35ꎬ56ꎬ36)ꎮ 合作用(4.23%ꎬ0ꎬ44)ꎮ 以上研究表明都匀地区
都匀地区茶独特的风味 是 由 叶 片 中 的 茶 氨 福鼎大白种茶独特的香味和各种健康功能主要归
酸、萜烯类和类黄酮 的 代 谢 产 物 赋 予 的ꎮ Wu et 因于萜类物质合成、光合作用、 类黄酮的生物合成
al.(2013)从茶树叶片的转录组研究中得出大多数 等生物过程ꎬ类黄酮主要赋予茶的苦味ꎬ二萜类化
基因参与编码合成黄酮类、咖啡碱和茶氨酸等次 合物的水解产物具有令人愉悦的香味和特色风
生代谢途径的重要酶ꎮ 该文通过 KEGG 功能显著 味ꎮ 为进一步研究都匀地区福鼎大白种茶生长发
性富集分析ꎬ发现叶和茎、叶和根、茎和根差异基 育和遗传育种提供了分子基础ꎬ并为揭示与类黄
因参与的部分极其显著的通路有类胡萝卜素生物 酮相关代谢通路有关基因提供了重要理论依据ꎮ
合成ꎬ泛醌和其他萜烯类醌的生物合成ꎬ单萜生物
合成ꎬ甘氨酸、丝氨酸和苏氨酸代谢ꎬ乙醛酸和二
参考文献:
羧酸代谢ꎬ植物激素信号转导ꎬ卟啉和叶绿素代
谢ꎬ苯丙素的生物合成ꎬ光合作用天线蛋白和光合 ANDERS Sꎬ REYES Aꎬ HUBER Wꎬ 2012. Detecting
differential usage of exons from RNA ̄seq data[J]. GenoResꎬ
作用ꎬ类黄酮生物合成ꎬ萜类化合物的生物合成ꎬ
22(10): 2008-2017.
亚麻酸代谢等显著富集ꎮ 叶和茎的差异表达基因 CHEN LBꎬ XIA LFꎬ ZHOU Mꎬ et al.ꎬ 2015. Analysis on tran ̄