
Page 157 - 《广西植物》2025年第3期
P. 157
3 期 姚淑婷等: 六堡茶群体种叶绿体基因组捕获历史与遗传多样性研究 5 3 7
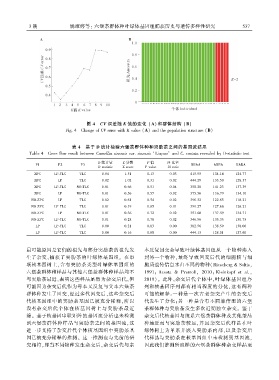
图 4 CV 误差随 K 值的变化 (A)和群体结构 (B)
Fig. 4 Change of CV error with K value (A) and the population structure (B)
表 4 基于 D 统计检验六堡茶群体种和突肋茶之间的基因流结果
Table 4 Gene flow result between Camellia sinensis var. sinensis ‘Liupao’ and C. costata revealed by D ̄statistic test
D 统计量 Z 分数 P 值 f4 比率
P1 P2 P3 BBAA ABBA BABA
D statistic Z score P value f4 ratio
ZPC LP ̄TLC TLC 0.04 1.51 0.13 0.03 415.55 134.14 124.77
ZPC LP TLC 0.02 1.02 0.31 0.02 444.29 133.50 128.37
ZPC LP ̄TLC NO ̄TLC 0.01 0.66 0.51 0.04 358.28 141.23 137.39
ZPC LP NO ̄TLC 0.01 0.56 0.57 0.02 375.56 136.79 134.10
NO ̄ZPC LP TLC 0.02 0.61 0.54 0.02 396.32 122.65 118.21
NO ̄ZPC LP ̄TLC TLC 0.01 0.19 0.85 0.01 399.27 127.66 126.21
NO ̄ZPC LP NO ̄TLC 0.01 0.36 0.72 0.02 351.60 137.59 134.71
NO ̄ZPC LP ̄TLC NO ̄TLC 0.01 0.28 0.78 0.02 346.56 133.35 130.75
LP LP ̄TLC TLC 0.00 0.21 0.83 0.00 382.56 138.59 138.06
LP LP ̄TLC TLC 0.00 0.16 0.88 0.00 444.13 128.01 127.60
最可能原因是它们的祖先与部分突肋茶的祖先发 本反复回交会导致叶绿体基因组从一个物种渗入
生了杂交ꎬ捕获了突肋茶的叶绿体基因组ꎮ 在串 到另一个物种ꎬ最终导致回交后代的细胞核与细
联核基因树上ꎬ含有突肋茶类型叶绿体基因组的 胞质遗传信息来自不同的物种(Rieseberg & Soltisꎬ
六堡茶群体种样品与其他六堡茶群体种样品均不 1991ꎻ Acosta & Premoliꎬ 2010ꎻ Kleinkopf et al.ꎬ
与突肋茶最近ꎬ表明这些样品虽然为杂交后代ꎬ但 2019)ꎮ 此外ꎬ杂交后代个体中ꎬ叶绿体基因组序
可能因为杂交后代作为母本又反复与父本六堡茶 列和核基因序列都有相当程度的分化ꎬ这有两种
群体种发生了回交ꎬ经过多代回交后ꎬ这些杂交后 可能的解释:一种是一次古老杂交产生的杂交后
代核基因组中的突肋茶基因已被充分稀释ꎬ所以 代发生了分化ꎻ另一种是含有不同单倍型的六堡
没有杂交后代个体在核基因树上与突肋茶最近 茶群体种与突肋茶发生多次近期独立杂交ꎮ 鉴于
缘ꎮ 基于核基因串联矩阵的基因流分析也未检测 杂交后代核基因与纯系六堡茶群体种及其他茶品
到六堡茶群体种样品与突肋茶之间的基因流ꎬ这 种最近而与突肋茶较远ꎬ并且杂交后代样品在叶
进一步支持了杂交后代个体核基因组中突肋茶基 绿体树上为单系并嵌入突肋茶内部ꎬ以及杂交后
因已被充分稀释的推测ꎮ 这一推测也与先前的研 代样品与突肋茶在核基因组中未检测到基因流ꎬ
究相符ꎬ即当不同物种发生杂交后ꎬ杂交后代与亲 因此我们推测所取样的六堡茶群体种杂交样品更